Corona “vaccination” – the body’s programmed self-destruction – How mRNA “vaccination” makes the immune system attack its own body
By
FRONT NEWS and additions by Robert Gorter, MD, PhD.
October 31st, 2021
http://robert-gorter.info/aerosols-transmit-prions-discussion-robert-gorter-md-phd/
http://robert-gorter.info/rnas-behave-like-prions/
‘Friendly fire.’ It is only thanks to our immune system that we can live in a world full of bacteria, viruses, parasites, and other pathogens in us and around us. It effectively protects us from germs. Once a germ or foreign cell enters our body, the immune system recognizes it based on features on the cell surface called antigens and destroys the cell carrying those antigens, wrote Dr. Christian Fiala and Dr. Robert Gorter.
This principle is used in vaccinations. For this purpose, a pathogen is usually weakened or killed so that it can no longer cause disease. However, its features, the antigens, remain on the cell surface. So the weakened or killed pathogen can still be recognized as foreign by the immune system when introduced into the body. This leads to the destruction of the pathogen, or the cell is recognized as foreign and destroyed.
An essential characteristic of the immune system is therefore the strict differentiation between its own (autologous) and foreign (heterologous) cells. Only in this way is healthy living possible at all. It is precisely this fundamental distinction and the basis of life that is being turned upside down by the injection, which is currently promoted as a corona “vaccination”. The much-cited spike proteins are a recognition feature of the Coronavirus, an antigen. When a virus enters the body, the immune system recognizes the invader based on the antigen and destroys the cells already infected by the virus.
With the mRNA corona “vaccination”, body cells are purposefully and for the first time in history genetically programmed to present the spike protein as a (foreign) antigen on their cell surface, even though they are perfectly healthy and not infected with the virus. The mechanism of action of the corona “vaccination” is to falsely mark one’s own healthy cells as foreign. The immune system reacts immediately, recognizing the antigen as foreign, forming antibodies against it, and destroying the spike protein by destroying the cell carrying this spike protein.
This means that the corona “vaccination” fools our immune system into attacking and destroying our own healthy cells. In the military, this is called ‘friendly fire’ when soldiers attack their own troops.
However, this abolition of the strict boundary between the foreign and the self poses a fundamental danger, not only to our health but also to our very survival. We are familiar with this situation with rare autoimmune diseases. These are usually serious and can even be fatal.
The dose makes the poison
In order to better estimate the danger of the Corona “vaccination”, it is important to know how much mRNA is administered with one dose of the vaccine, or how many body cells are stimulated to produce the spike protein and thus be released to be destroyed by the immune system. Unfortunately, no information from the manufacturers is available on this, nor is this important information mentioned in the approval studies. There is only a scientific estimate of the number of particles in which the mRNA is transported, the so-called lipid nanoparticles. By this estimate, one injection contains a staggering number of approximately 2 trillion particles. That is written out as 2,000,000,000,000 or 2,000 billion parts. Now one can assume that not every particle reaches a healthy cell, and that some cells in the body are affected by more than one particle. In addition, not all particles may be fully functional.
More and more doctors against COVID-19 vaccination of children
Nevertheless, this unimaginably large number of particles, which are programmed to destroy healthy body cells, is extremely relevant when one considers that the human body consists of approximately 37 trillion cells. In view of the incalculable risks of this new technology, it is important to note that vaccination against the new coronavirus can also be achieved with the existing technology with an attenuated pathogen.
What organs does the Corona “vaccination” damage?
The Corona vaccine is programmed in such a way that the immune system destroys its own cells that produce the spike protein and present it to the cell surface. But in which organs does this happen? The answer to this question can be found in animal experiments by the vaccine manufacturer BioNTech/Pfizer. The blueprint for the spike protein can be found in a gene segment, the so-called mRNA. However, because gene segments are very fragile and break easily outside a cell, they are embedded in very small fat globules called lipid nanoparticles.
After injection, these quickly pass into the blood and then into the body cells, so that after 1 hour half of them have already been distributed throughout the body. In the above animal experiment, the lipid nanoparticles were found in all organs, as expected especially in the liver. But they were also discovered in the brain, proving that they were able to cross the highly effective blood-brain barrier. They have also been found in the ovaries and testes, which also prove that they can cross the testicular-blood barrier.
According to the purely random distribution of lipid nanoparticles throughout the body, the body cells from the spike protein in the respective organs. This could explain why the very numerous side effects and vaccine damage are also found randomly distributed in many organs and depending on their severity they can also be fatal.
Vaccinated people are carriers of more infectious and antibody-resistant variants
Since the lipid nanoparticles are distributed with the blood throughout the body, the cells that line the blood vessels, the so-called endothelial cells, are particularly affected. This explains the numerous blood clots (thrombosis or emboli) observed as a result of the corona “vaccination”. These also occur in the brain, for example, and lead to partially irreversible damage there. This is particularly disturbing, not only because the brain is the central organ, but also because it shows that even organs that are separated from the immune system by a special barrier, such as the brain and testicles, are damaged by the corona “vaccination”. . The undirected and purely random distribution of the mRNA in the body and thus the random distribution of the destruction of somatic cells brought about by the Corona “vaccination” can be compared to a shotgun shot in a flock of birds. You don’t know how many birds you’ll hit, but there’s a good chance you’ll kill a few.
Booster “vaccination” – enough is not enough
Unlike most other vaccinations, the corona “vaccination” must be approved by the government or health authorities at short intervals. For example, the regulations of the Minister of Health state that the vaccination loses its validity after 270 days (9 months), and the vaccinated persons are then regarded as unvaccinated. In addition, the USA government has already ordered 40 million vaccinations for the next two years (2022 and 2023) for 9 million inhabitants. That is why a booster vaccination has now been announced for autumn 2021 for everyone who has already been fully vaccinated. In other countries, such as Israel, a large part of the population has already been vaccinated for the third time. The question therefore arises what effect a repeated injection has on the body.
Our immune system is able to learn. When a pathogen first enters the body, it is immediately recognized as foreign, but the formation of defense measures and the destruction of the pathogen take several days. During this time, the pathogen prevails for a short time, which is why we get sick. Only after a few days the immune system is strong enough to destroy the pathogen and we become healthy. Fortunately, the immune system remembers the “appearance” of the pathogen and its antigenic properties of the pathogen and when it comes into contact with it again, it is activated much faster and much stronger.
Ten Legitimate Reasons Why So Many People Refuse Covid Injections
This vital mechanism also plays out with each subsequent Corona “vaccination”. However, the massive response of the immune system with each new injection is not directed against a pathogen, but again against our own healthy body cells. Because of the marking with the spike protein, the immune system sees the body cells again as a pathogen that must be destroyed.
But on the second injection and on all subsequent injections, he is better prepared and much stronger. This means that with all subsequent injections, the body cells are destroyed even more efficiently by the immune system than with the first injection. Thus, this memory capacity of the immune system, which is important for our survival, becomes a dangerous weapon against ourselves, because with each repeated injection it turns against our own body cells with each repeated injection. Instead of making foreign pathogens harmless, it destroys our own cells that produce the spike protein.
Any further injection of a Corona “vaccination” therefore carries a greater and greater risk. As with the first injection, all further injections destroy healthy body cells,
But to a much greater extent, because the immune system is primed and therefore able to destroy cells producing the spike protein much more efficiently. This is consistent with the observation that more and stronger vaccination complications occur with the second vaccination. The same applies to people who have had a corona infection in the past, have recovered, and are still receiving a corona vaccination. Sticking to the shotgun and flock of birds example above, if you shoot a flock of birds a few times, you won’t know what you would hit, but with each new shot, the number of birds gets smaller – until finally no flock of birds more. This mode of action also means that people who have already been vaccinated.
Studies with various mRNA vaccines in the large animals’ University of Utrecht, The Netherlands: RNAs That Behave Like Prions
Messenger RNA Definition
Messenger ribonucleic acids (mRNAs) transfer the information from DNA to the cell machinery that makes proteins. Tightly packed into every cell nucleus, which measures just 10 microns in diameter, is a three-meter long double-stranded DNA “instruction manual” on how to build and maintain a human body. In order for each cell to maintain its structure and perform all of its functions, it must continuously manufacture cell-type-specific parts (proteins). Inside each nucleus, a multi-subunit protein called RNA polymerase II (RNAP II) reads DNA and simultaneously fabricates a “message” or transcript, which is called messenger RNA (mRNA), in a process called transcription. Molecules of mRNA are composed of relatively short, single strands of molecules made up of adenine, cytosine, guanine, and uracil bases held together by a sugar-phosphate backbone. When RNA polymerase finishes reading a section of the DNA, the pre-mRNA copy is processed to form mature mRNA and then transferred out of the cell nucleus. Ribosomes read the mRNA and translate the message into functional proteins in a process called translation. Depending on the newly synthesized protein’s structure and function, it will be further modified by the cell, exported to the extracellular space, or will remain inside the cell. The diagram below shows transcription (DNA->RNA) taking place in the cell nucleus where RNAP is RNA polymerase II enzyme synthesizing RNA.
Functions of mRNA
The primary function of mRNA is to act as an intermediary between the genetic information in DNA and the amino acid sequence of proteins. mRNA contains codons that are complementary to the sequence of nucleotides on the template DNA and direct the formation of amino acids through the action of ribosomes and tRNA. mRNA also contains multiple regulatory regions that can determine the timing and rate of translation. In addition, it ensures that translation proceeds in an orderly fashion because it contains sites for the docking of ribosomes, tRNA as well as various helper proteins.
Proteins produced by the cells play a variety of roles, either as enzymes, structural molecules or transport machinery for various cellular components. Some cells are also specialized for secreting proteins, such as the glands that produce digestive enzymes or hormones which influence the metabolism of the entire organism.
How do mRNA vaccinations work?
In vaccinations with mRNA, the idea is to have the synthesized mRNA initiate the production of COVID-19 spike proteins being in ribosomes and through this pathway, the host cell would infect itself with self-made spike protein. Natural mRNA carries information from the DNA in the nucleus to the ribosomes as a messenger.
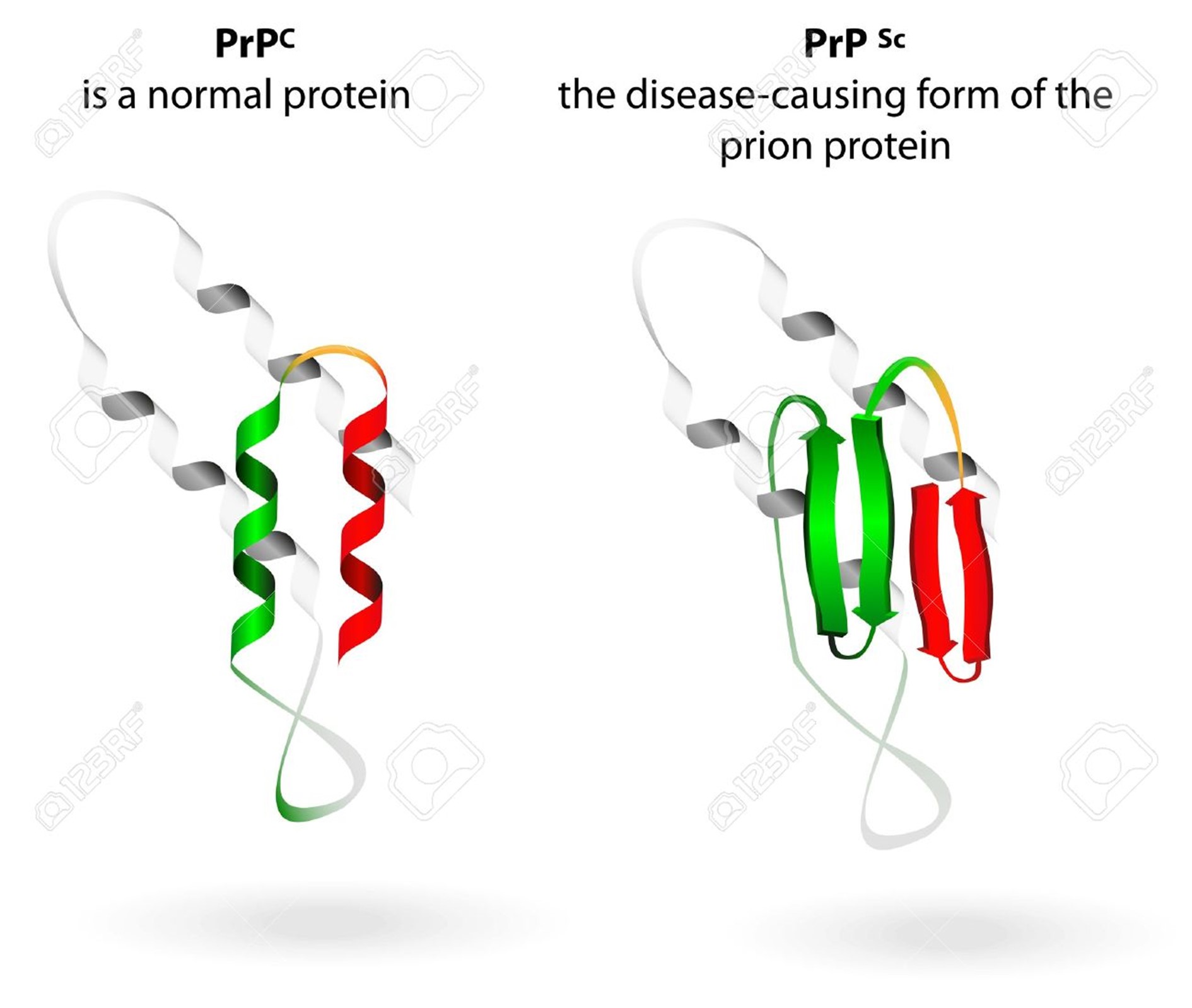
What Are Prions?
Prions are distorted versions of normal proteins found in the human and animal brains and other tissues. These distorted (‘misfolded’) proteins damage brain cells, leading to fatal dementias akin to human Alzheimer’s and Parkinson’s diseases. One of the amazing things about prions is that, although they are composed only of proteins and contain no genes more related to RNA-like molecules. They have acquired the ability to be transmitted like any other infectious agent such as viruses or bacteria. Best known prion diseases are mad cow disease of cattle and humans, Creutzfeld-Jakob disease of humans, chronic wasting disease of deer and elk, and scrapie of sheep. A better understanding of prions is important in controlling the threat they represent to animals and humans and for what we can learn about the cause and treatment of the major dementia diseases of humans.
Humans are usually infected by eating infected raw meat (“Beefsteak Tartare” and undercooked meat products) and prions can be found in blood and can transmit by blood transfusions and blood products.
Current injections with mRNA contain approx. 70% purified mRNA to inform ribosomes to ensemble COVID-19 spike proteins. What the other 30% mRNA molecules will cause ribosomes to create is unknown but likely aberrant proteins, like prions.
Ribosomes are enzymes composed of many proteins that catalyze the synthesis of proteins from mRNA in the process of translation. Ribosomes exist freely in the cell cytoplasm or remain attached to the endoplasmic reticulum.
Conclusion
The real fall of humanity is not the desire to acquire knowledge, but through knowledge in the hands of criminal psychopaths.
To abolish the essential division between foreign pathogens and the body’s own cells. With the mRNA-corona “vaccination” an unimaginably large number of healthy endogenous cells with the spike protein is specified as foreign and dangerous and thus, released for destruction by the own immune system.
And then the effects of prions are still not completely known but in large animal studies at the University of Utrecht, The Netherlands, (2000-2013) but all had very bad results, showing massive deaths due to autoimmune and prion disease.

References
http://robert-gorter.info/aerosols-transmit-prions-discussion-robert-gorter-md-phd/
http://robert-gorter.info/rnas-behave-like-prions/
1.↵Alper T. 1972. The nature of the scrapie agent. J Clin Pathol Suppl (R Coll Pathol) 25:154–155. doi:10.1136/jcp.25.Suppl_6.154.FREE Full TextGoogle Scholar
2.↵Bolton DC, McKinley MP, Prusiner SB. 1982. Identification of a protein that purifies with the scrapie prion. Science 218:1309–1311. doi:10.1126/science.6815801.Abstract/FREE Full TextGoogle Scholar
3.↵Diringer H, Gelderblom H, Hilmert H, Özel M, Edelbluth C, Kimberlin RH. 1983. Scrapie infectivity, fibrils and low molecular weight protein. Nature 306:476–478. doi:10.1038/306476a0.CrossRefPubMedGoogle Scholar
4.↵Diener TO, McKinley MP, Prusiner SB. 1982. Viroids and prions. Proc Natl Acad Sci U S A 79:5220–5224. doi:10.1073/pnas.79.17.5220.Abstract/FREE Full TextGoogle Scholar
5.↵Bellinger-Kawahara C, Diener TO, McKinley MP, Groth DF, Smith DR, Prusiner SB. 1987. Purified scrapie prions resist inactivation by procedures that hydrolyze, modify, or shear nucleic acids. Virology 160:271–274. doi:10.1016/0042-6822(87)90072-9.CrossRefPubMedWeb of ScienceGoogle Scholar
6.↵Griffith JS. 1967. Nature of the scrapie agent: self-replication and scrapie. Nature 215:1043–1044. doi:10.1038/2151043a0.CrossRefPubMedWeb of ScienceGoogle Scholar
7.↵Dickinson AG, Meikle VM, Fraser H. 1968. Identification of a gene which controls the incubation period of some strains of scrapie agent in mice. J Comp Pathol 78:293–299. doi:10.1016/0021-9975(68)90005-4.CrossRefPubMedWeb of ScienceGoogle Scholar
8.↵Scott M, Foster D, Mirenda C, Serban D, Coufal F, Wälchli M, Torchia M, Groth D, Carlson G, DeArmond SJ, Westaway D, Prusiner SB. 1989. Transgenic mice expressing hamster prion protein produce species-specific scrapie infectivity and amyloid plaques. Cell 59:847–857. doi:10.1016/0092-8674(89)90608-9.CrossRefPubMedWeb of ScienceGoogle Scholar
9.↵Zabel MD, Reid C. 2015. A brief history of prions. Pathog Dis 73:ftv087. doi:10.1093/femspd/ftv087.CrossRefPubMedGoogle Scholar
10.↵Aigle M, Lacroute F. 1975. Genetical aspects of [URE3], a non-mitochondrial, cytoplasmically inherited mutation in yeast. Mol Gen Genet 136:327–335. doi:10.1007/BF00341717.CrossRefPubMedWeb of ScienceGoogle Scholar
11.↵Wickner RB. 1994. [URE3] as an altered URE2 protein: evidence for a prion analog in Saccharomyces cerevisiae. Science 264:566–569. doi:10.1126/science.7909170.Abstract/FREE Full TextGoogle Scholar
12.↵Taylor KL, Cheng N, Williams RW, Steven AC, Wickner RB. 1999. Prion domain initiation of amyloid formation in vitro from native Ure2p. Science 283:1339–1343. doi:10.1126/science.283.5406.1339.Abstract/FREE Full TextGoogle Scholar
13.↵Speransky VV, Taylor KL, Edskes HK, Wickner RB, Steven AC. 2001. Prion filament networks in [Ure3] cells of Saccharomyces cerevisiae. J Cell Biol 153:1327–1336. doi:10.1083/jcb.153.6.1327.Abstract/FREE Full TextGoogle Scholar
14.↵Kushnirov VV, Vishnevskaya AB, Alexandrov IM, Ter-Avanesyan MD. 2007. Prion and nonprion amyloids: a comparison inspired by the yeast Sup35 protein. Prion 1:179–184. doi:10.4161/pri.1.3.4840.CrossRefPubMedWeb of ScienceGoogle Scholar
15.↵Saupe SJ. 2011. The [Het-s] prion of Podospora anserina and its role in heterokaryon incompatibility. Semin Cell Dev Biol 22:460–468. doi:10.1016/j.semcdb.2011.02.019.CrossRefPubMedGoogle Scholar
16.↵Wickner RB, Edskes HK, Ross ED, Pierce MM, Baxa U, Brachmann A, Shewmaker F. 2004. Prion genetics: new rules for a new kind of gene. Annu Rev Genet 38:681–707. doi:10.1146/annurev.genet.38.072902.092200.CrossRefPubMedWeb of ScienceGoogle Scholar
17.↵Halfmann R, Jarosz DF, Jones SK, Chang A, Lancaster AK, Lindquist S. 2012. Prions are a common mechanism for phenotypic inheritance in wild yeasts. Nature 482:363–368. doi:10.1038/nature10875.CrossRefPubMedWeb of ScienceGoogle Scholar
18.↵Holmes DL, Lancaster AK, Lindquist S, Halfmann R. 2013. Heritable remodeling of yeast multicellularity by an environmentally responsive prion. Cell 153:153–165. doi:10.1016/j.cell.2013.02.026.CrossRefPubMedWeb of ScienceGoogle Scholar
19.↵Jarosz DF, Brown JCS, Walker GA, Datta MS, Ung WL, Lancaster AK, Rotem A, Chang A, Newby GA, Weitz DA, Bisson LA, Lindquist S. 2014. Cross-kingdom chemical communication drives a heritable, mutually beneficial prion-based transformation of metabolism. Cell 158:1083–1093. doi:10.1016/j.cell.2014.07.025.CrossRefPubMedGoogle Scholar
20.↵Wickner RB, Shewmaker FP, Bateman DA, Edskes HK, Gorkovskiy A, Dayani Y, Bezsonov EE. 2015. Yeast prions: structure, biology, and prion-handling systems. Microbiol Mol Biol Rev 79:1–17. doi:10.1128/MMBR.00041-14.Abstract/FREE Full TextGoogle Scholar
21.↵Manjrekar J. 2017. Epigenetic inheritance, prions and evolution. J Genet 96:445–456. doi:10.1007/s12041-017-0798-3.CrossRefGoogle Scholar
22.↵Wickner RB, Edskes HK, Roberts BT, Baxa U, Pierce MM, Ross ED, Brachmann A. 2004. Prions: proteins as genes and infectious entities. Genes Dev 18:470–485. doi:10.1101/gad.1177104.FREE Full TextGoogle Scholar
23.↵Roberts BT, Wickner RB. 2003. Heritable activity: a prion that propagates by covalent autoactivation. Genes Dev 17:2083–2087. doi:10.1101/gad.1115803.Abstract/FREE Full TextGoogle Scholar
24.↵Roberts BT, Wickner RB. 2004. A new kind of prion: a modified protein necessary for its own modification. Cell Cycle 3:100–103. doi:10.4161/cc.3.2.642.CrossRefPubMedGoogle Scholar
25.↵Cancellotti E, Mahal SP, Somerville R, Diack A, Brown D, Piccardo P, Weissmann C, Manson JC. 2013. Post-translational changes to PrP alter transmissible spongiform encephalopathy strain properties. EMBO J 32:756–769. doi:10.1038/emboj.2013.6.Abstract/FREE Full TextGoogle Scholar
26.↵Chakravarty AK, Smejkal T, Itakura AK, Garcia DM, Jarosz DF. 2020. A non-amyloid prion particle that activates a heritable gene expression program. Mol Cell 77:251–265.e9. doi:10.1016/j.molcel.2019.10.028.CrossRefGoogle Scholar
27.↵Harvey ZH, Chakravarty AK, Futia RA, Jarosz DF. 2020. A prion epigenetic switch establishes an active chromatin state. Cell 180:928–‐940.e14. doi:10.1016/j.cell.2020.02.014.CrossRefGoogle Scholar
28.↵Badelt S, Flamm C, Hofacker IL. 2016. Computational design of a circular RNA with prionlike behavior. Artif Life 22:172–184. doi:10.1162/ARTL_a_00197.CrossRefGoogle Scholar
29.↵Ferré-D’Amaré AR, Scott WG. 2010. Small self-cleaving ribozymes. Cold Spring Harb Perspect Biol 2:a003574. doi:10.1101/cshperspect.a003574.Abstract/FREE Full TextGoogle Scholar
30.↵Ramesh A, Winkler WC. 2014. Metabolite-binding ribozymes. Biochim Biophys Acta 1839:989–994. doi:10.1016/j.bbagrm.2014.04.015.CrossRefGoogle Scholar
31.↵Ferré-D’Amaré AR. 2011. Use of a coenzyme by the glmS ribozyme-riboswitch suggests primordial expansion of RNA chemistry by small molecules. Philos Trans R Soc Lond B Biol Sci 366:2942–2948. doi:10.1098/rstb.2011.0131.CrossRefPubMedGoogle Scholar
32.↵Tinsley RA, Furchak JRW, Walter NG. 2007. Trans-acting glmS catalytic riboswitch: locked and loaded. RNA 13:468–477. doi:10.1261/rna.341807.Abstract/FREE Full TextGoogle Scholar
33.↵Lau MWL, Ferré-D’Amaré AR. 2013. An in vitro evolved glmS ribozyme has the wild-type fold but loses coenzyme dependence. Nat Chem Biol 9:805–810. doi:10.1038/nchembio.1360.CrossRefGoogle Scholar
34.↵Diener TO. 2003. Discovering viroids—a personal perspective. Nat Rev Microbiol 1:75–80. doi:10.1038/nrmicro736.CrossRefPubMedWeb of ScienceGoogle Scholar
35.↵Elena SF, Dopazo J, Flores R, Diener TO, Moya A. 1991. Phylogeny of viroids, viroidlike satellite RNAs, and the viroidlike domain of hepatitis delta virus RNA. Proc Natl Acad Sci U S A 88:5631–5634. doi:10.1073/pnas.88.13.5631.Abstract/FREE Full TextGoogle Scholar
36.↵Flores R, Grubb D, Elleuch A, Nohales MÁ, Delgado S, Gago S. 2011. Rolling-circle replication of viroids, viroid-like satellite RNAs and hepatitis delta virus: variations on a theme. RNA Biol 8:200–‐206. doi:10.4161/rna.8.2.14238.CrossRefPubMedWeb of ScienceGoogle Scholar
37.↵Daròs JA, Flores R. 1995. Identification of a retroviroid-like element from plants. Proc Natl Acad Sci U S A 92:6856–6860. doi:10.1073/pnas.92.15.6856.Abstract/FREE Full TextGoogle Scholar
38.↵Vera A, Daròs JA, Flores R, Hernández C. 2000. The DNA of a plant retroviroid-like element is fused to different sites in the genome of a plant pararetrovirus and shows multiple forms with sequence deletions. J Virol 74:10390–10400. doi:10.1128/jvi.74.22.10390-10400.2000.Abstract/FREE Full TextGoogle Scholar
39.↵Hegedus K, Dallmann G, Balázs E. 2004. The DNA form of a retroviroid-like element is involved in recombination events with itself and with the plant genome. Virology 325:277–286. doi:10.1016/j.virol.2004.04.035.CrossRefPubMedWeb of ScienceGoogle Scholar
40.↵Hammann C, Luptak A, Perreault J, de la Peña M. 2012. The ubiquitous hammerhead ribozyme. RNA 18:871–885. doi:10.1261/rna.031401.111.Abstract/FREE Full TextGoogle Scholar
41.↵Bussière F, Ledû S, Girard M, Héroux M, Perreault J-P, Matton DP. 2003. Development of an efficient cis-trans-cis ribozyme cassette to inactivate plant genes: cis-trans-cis self-cleaving ribozyme cassette. Plant Biotechnol J 1:423–435. doi:10.1046/j.1467-7652.2003.00039.x.CrossRefPubMedGoogle Scholar
42.↵Webb C-HT, Lupták A. 2018. Kinetic parameters of trans scission by extended HDV-like ribozymes and the prospect for the discovery of genomic trans-cleaving RNAs. Biochemistry 57:1440–1450. doi:10.1021/acs.biochem.7b00789.CrossRefGoogle Scholar
43.↵Huang X, Zhao Y, Pu Q, Liu G, Peng Y, Wang F, Chen G, Sun M, Du F, Dong J, Cui X, Tang Z, Mo X. 2019. Intracellular selection of trans-cleaving hammerhead ribozymes. Nucleic Acids Res 47:2514–2522. doi:10.1093/nar/gkz018.CrossRefGoogle Scholar
44.↵Czech B, Hannon GJ. 2016. One loop to rule them all: the ping-pong cycle and piRNA-guided silencing. Trends Biochem Sci 41:324–337. doi:10.1016/j.tibs.2015.12.008.CrossRefPubMedGoogle Scholar
45.↵Czech B, Munafò M, Ciabrelli F, Eastwood EL, Fabry MH, Kneuss E, Hannon GJ. 2018. piRNA-guided genome defense: from biogenesis to silencing. Annu Rev Genet 52:131–157. doi:10.1146/annurev-genet-120417-031441.CrossRefGoogle Scholar
46.↵Rechavi O, Lev I. 2017. Principles of transgenerational small RNA inheritance in Caenorhabditis elegans. Curr Biol 27:R720–R730. doi:10.1016/j.cub.2017.05.043.CrossRefGoogle Scholar
47.↵Battaglia E. 2005. Prion, proteon, proteo-conformer: a terminological analysis. Rend Fis Acc Lincei 16:5–17. doi:10.1007/BF02904737.CrossRefGoogle Scholar
48.↵de la Peña M. 2018. Circular RNAs biogenesis in eukaryotes through self-cleaving hammerhead ribozymes, p 53–63. In Xiao J (ed), Circular RNAs. Springer Singapore, Singapore.Google Scholar